
Die rol van koolhidrate in alternerende drag by pekanneutbome
Deur Dr. Elmi Lötze, Hoof van ITEST™CARBOHYDRATES en ITEST™LEAF en Deon Claassen, Senior Hortoloog
Alternerende drag
Koolhidrate speel ’n belangrike rol in die alternerende drag (of “oorslaan”) van baie vrugtesoorte, insluitend pekanneutbome.
Alternerende drag verwys na die verskynsel waar bome ’n groot oes in een jaar (die sogenaamde aan-jaar) produseer en ’n lae oes in die daaropvolgende jaar (af-jaar).
Die voorkoms van alternerende drag wissel en kan ’n tak, boom of boord behels (Monselise en Goldschmidt, 1982). Dit volg ook nie noodwendig ’n twee-jaar siklus nie, waar twee tot drie aan- of af-jare na mekaar kan voorkom (Wood et al., 2003).
Alternerende drag is wêreldwyd die belangrikste ekonomiese uitdaging in pekanneutproduksie (Wood et al., 2003). Enige manipulasie om die voorkoms daarvan te verminder of geheel en al te voorkom, sal dus ’n reuse bydrae maak tot die suksesvolle (en ekonomiese) verbouing van pekanneute.
’n Metode om alternerende drag te kwantifiseer is deur die berekening van ’n alternerende indeks, wat die intensiteit van alternerende drag bepaal. Die intensiteit wissel van 0, waar opbrengste min of meer konstant bly tussen jare, tot 1, waar totale alternering voorkom in ’n jaar met geen opbrengs nie (Conner en Worley, 2000).

Figuur 1: Waterblasiestadium van pekanneute in Maart.
Alternatiewelik sal ’n akkurate oesvoorspellingsmodel nie alleen die kans op alternering voorspel nie, maar ook die toe- of afname in produksie kwantifiseer en so verdere beplanning ondersteun.
Faktore wat alternering bepaal
Alternerende drag word gereguleer deur voeding, hormone en die koolhidraatstatus van die boom. Die voorkoms van alternerende drag by pekanneutbome is hoofsaaklik te wyte aan drie faktore wat verband hou met energie: die lang tydperk wat dit neem vir neute om volwassenheid te bereik, vruggroei/vergroting en die chemiese samestelling van die neutkern (Sparks, 1974).
Koolhidrate word geassosieer met die meeste fenologiese stadia gedurende die ontwikkeling van die neut – vanaf blominduksie tot oes.
Volgens die Koolhidraat-teorie word blom- en vrugvorming direk bepaal deur die koolhidraatreserwes wat tydens die dormante periode opbou (Barnet en Mielke, 1981).
Daarteenoor steun die Fitohormoon-teorie op hormonale beheer van die prosesse via vrugte, blare of beide. Verder is daar ’n noue verwantskap tussen die vervoer van hormone en suiker, en die hormonale sein vir biochemiese reaksies. Blominduksie, as voorbeeld, moet ondersteun word deur energie afkomstig van koolhidrate vir die finale uitkoms, byvoorbeeld ’n reproduktiewe knop.
Blomontwikkeling en vruggroei (neut) is tot die koolhidraatreserwes wat in die dormante seisoen opgebou het. Kultivars wat vroeg volwassenheid bereik, het dus ’n langer periode om koolhidrate op te bou en is minder onderhewig aan alternering as laat-kultivars.
Die neutkern akkumuleer ongeveer tweederdes van sy totale droë gewig gedurende die 80 dae voor blaarval en bevat ’n hoë persentasie lipiede (≈ 70%). Dit beteken dat ’n groot hoeveelheid energie gemobiliseer moet word in die kort periode voor blaarval ten einde in hierdie behoefte te voorsien (Monselise en Goldschmidt, 1982). Koolhidraatreserwes moet dus betyds aangevul word, voor én na hierdie tydperk, vir beide neutvul asook dormansie daarna.
Koolhidraatbestuur en alternerende drag
Tydens ’n aan-jaar word ’n groot deel van die koolhidrate wat deur fotosintese geproduseer word, verbruik vir neutontwikkeling en -groei.
Die gevolg hiervan is dat die koolhidraatreserwes in die boom, veral stysel, aansienlik verminder. Dit kan lei tot ’n afname in vegetatiewe groei, en blomknopvorming in die daaropvolgende jaar inhibeer. In die af-jaar met ’n verminderde oeslading, het die boom ’n oormaat koolhidrate. Dit het weer die akkumulasie van styselreserwes tot gevolg, wat vegetatiewe groei en die vorming van blomknoppe vir die volgende jaar se oes bevorder.
Fotosintese
Die grootte van die aktiewe energie- en reserwepoel hou verband met die fotosintetiese kapasiteit van die blare tydens die seisoen (Conner en Worley, 2000). Dus is die eerste prioriteit van die produsent om ’n gesonde blaaroppervlak met voldoende ligonderskepping en -verspreiding in die boord te verseker. Aangesien water en minerale elemente ook vir fotosintese benodig word, is besproeiingskedulering en ’n gefokusde bemestingstrategie van kritieke belang. Al hierdie praktyke impakteer die fisiologiese status van die boom.
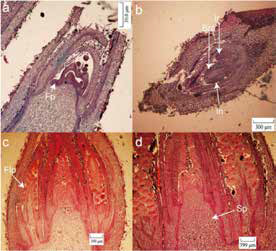
Figuur 2: Morfologiese eienskappe van vroeë manlike blomknopdifferensiasie van Mahan, China (Xie et al. 2023).
(a) Vroeë stadium van differensiasie met blomprimordia (Fp) laat Oktober (RSA).
(b) Manlike blomprimordia met omhulsel (In), skutblaar (Bra) en bloeiwyse rachis (Ir) vanaf middel Oktober.
(c) Blomprimordia (Flp) verskyn saam met die verlenging van die manlike bloeiwyse en toename in skutblare in die begin van Januarie.
(d) Die meeldraadprimordium (Sp) raak sigbaar na kelkprimordium-differensiasie einde Oktober.
Koolhidraatontledings
’n Uitstekende manier om hierdie status te kwantifiseer is om koolhidraatontledings op kritieke aksietye te neem en dan die resultaat te interpreteer in terme van die historiese inligting, om toekomstige bestuursaksies te bepaal. So kan die koolhidraatstatus van wortels en lote aan die einde van die dormante seisoen ’n aanduiding wees van i) die na-oes-strategie om koolhidraatreserwes aan te vul, ii) die benutting van reserwe koolhidrate gedurende die winter, en iii) wat verwag kan word in die komende seisoen in terme van beskikbare koolhidrate vir bot en die aanvanklike groei in die lente.
Lae styselvlakke in die weefsels gedurende hierdie stadium is ’n voortydige aanduiding dat fotosintese vroeg gestimuleer behoort te word en, in die geval waar ’n groot oes set, dat opvolg ondersteuning noodsaaklik gaan wees.
Om die hoeveelheid koolhidrate te verhoog vir die volgende jaar, kan verskillende manipulasies op plaasvlak toegepas word om optimale fotosintese en die vervoer en verspreiding van suikers te verseker, onder meer die bestuur van peste en plae, verhoogde sonligonderskepping en -verspreiding deur snoeipraktyke en plantdigtheid, goeie besproeiingskedulering en plantvoeding.
Voorts kan manipulasies soos ringelering, groeistremming en vrugset met plantgroeireguleerders ook ingespan word vir koolhidraaten/ of hormoonbestuur tydens set. Die effektiwiteit van die manipulasies varieer tussen kultivars, tyd van uitvoer en strafheid.
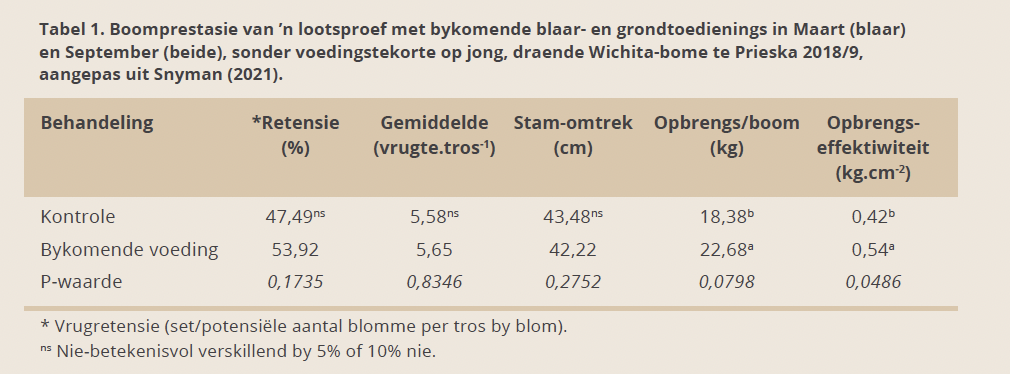
Lootsproef
In ’n belowende, plaaslike lootsproef, het ’n addisionele voedingsblaarspuit gedurende Maart (jaar 1 & 2) op ’n blok sonder voedingstekorte (blaaranalise), opgevolg met ’n grondbemesting/ biostimulant-kombinasie in September (jaar 2), gelei tot ’n betekenisvolle verhoging van opbrengs (P < 0,10) en opbrengseffektiwiteit (P < 0,05) op jong, (5 – 6 jaar) draende Wichitabome die daaropvolgende seisoen (jaar 2) (Tabel 1).
Die toediening in Maart (waterblasiestadium (Figuur 1) laat differensiasie stadium van manlike blomme van daaropvolgende jaar (Figuur 2) het moontlik ’n rol in fotosintese gespeel, wat die opbou van reserwes asook voedingselemente daarna kon ondersteun vir die tweede jaar se oes. Die aanvulling in September (knopswel (Figuur 3) differensiasie van vroulike blomme van volgende jaar (Figuur 4)) speel ’n rol in die bestuur van energie vir die oes in jaar 2.
Die morfologiese ontwikkeling van die manlike en vroulike blomme vir Mahan in China vir ongeveer September (RSA), word geïllustreer in Figure 1 en 2 (Xie et al., 2023).
Die effek van die behandelings op die koolhidraatstatus van die boom, en dus opbrengseffektiwiteit, is nie in die tesis gekwantifiseer nie. Agri Technovation beplan om die komende seisoen met ’n soortgelyke benadering te begin en dit te kwantifiseer met die ITEST™CARBOHYDRATE-diens op kommersiële boorde.
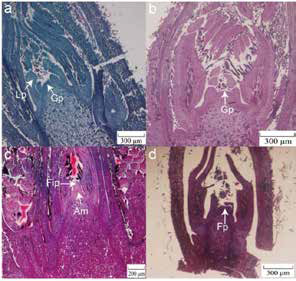
Figuur 4: Morfologiese eienskappe van vroulike blomdifferensiasie in Mahan, China (Xie et al. 2023).
(a) Die groeipunt (Gp) van die knop raak Kleiner en gepunt en die blaarprimordium (Lp) begin differensieer – begin November (RSA).
(b) Blomknoppe se groeipunt plat af en knoppe bereik die kritieke periode van differensiasie wat strek tot einde Desember.
(c) Vanaf laat Desember tot middel Januarie verleng knop groeipunte opwaarts en vorm ’n prominente sfeer. Die apikale meristeen
(Am) en vroulike bloeiwyse-primordia (Fip) word sigbaar.
(d) Na nog morfologiese veranderinge= differensieer die uitstulpings in vroulike primordia (Fp).
Verwysings:
- Barnett, J. and Mielke, E.A. 1981. Alternate bearing: A re-evaluation. Pecan South 8: 20-30.
- Conner, P.J. and Worley, R.E. 2000. Alternate bearing intensity of pecan cultivars. HortScience 35: 1067-1069.
- Monselise, S.P. and Goldschmidt, E.E. 1982. Alternate bearing in fruit trees. Horticultural Reviews 4: 128-173.
- Snyman, T.J. 2021. Investigating the reproductive potential and yield of Pecans. [Caryaillinoinensis (Wangenh. K. Koch)] under South African conditions. MSc Agric thesis, Stellenbosch University.
- Sparks, D. 1974. The alternate fruit bearing problem in pecans. Annual Report, Northern Nut Growers Association 47: 145-158.
- Wood et al. 2003. Relationship of alternate bearing intensity in pecan to fruit and canopy characteristics. HortScience 38: 361-366.
- Xie et al. 2023. Transcriptional regulation of female and male flower bud initiation and development in Pecan (Carya illinoensis). Plants 12(6), 1378.



